🌸 Male Sterility in Plants
Types of male sterility and their practical use in hybrid seed production.
This lesson covers core principles and exam-focused points from this topic in plant breeding.
Self-Incompatibility
Self-incompatibility and sterility are the two mechanisms, which encourage cross
pollination. More than 300 species belonging to 20 families of angiosperms show self
incompatibility.
Definition
In self incompatibility plants, the flowers will produce functional or viable pollen grains
which fail to fertilize the same flower or any other flower of the same plant.
-
Self incompatible pollen grain may fail to germinate on the stigmatic surface.
-
Some may germinate but fails to penetrate the stigmatic surface.
-
Some pollen grains may produce pollen tube, which enters through stigmatic surface, but
its growth will be too slow. By the time the pollen tube enters the ovule the flower will
drop.
- Some time fertilziation is effected but embryo degenerates early.
Reason
Self-incompatibility is appeared to be due to biochemical reaction, but precise nature of
these is not clearly understood.
Classification of self incompatibility
According to Lewis (1954) the self incompatibility is classified as follows:
Self incompatibility
| Col1 | Col2 |
|---|---|
Distyly Tristylty Gametophytic system Sporophytic system
Heteromorphic system
In this case there will be difference in the morphology of the flowers. For example in
Primula sp there are two types of flowers namely PIN and THRUM. PIN flowers have long style
and short stamens while THRUM flowers have short style and long stamens. This type of
difference is known as Distyly. In case of distyly the only compatible mating is between PIN and
THRUM. The relative position of anthers is determined by single gene S/s. The recessive as
produces PIN and heterozygotes Ss produces THRUM.
Homozygous dominant SS is lethal and do not exist. The incompatibility reaction of
pollen is determined by the genotype of the plant producing them. Allele S is dominant over s.
This system is also known as heteromorphic – sporophytic system. Pollen grains produced by
PIN flowers will be all s in genotype as well as in incompatibility reaction: Whereas THRUM
flowers will produce two types of gametes S and s but all of them would be S phenotypically.
The mating between PIN and THRUM would produce Ss and ss progeny in equal frequencies.
This system is of little importance in crop plants. It occurs in sweet potato and bulk wheat.
| PIN FLOWER | Col2 | THRUM FLOWER | Col4 |
|---|---|---|---|
| Mating | Mating | Progeny | Progeny |
| Phenotype | Genotype | Genotype | Phenotype |
| Pin x Pin | ss x ss | Incom. Mating | - |
| Pin x Thrum | SS x Ss | 1 ss : Ss | 1Thrum 1 Pin |
| Thrum x Pin | Ss x ss | 1 Ss : ss | 1 Thrum 1 Pin |
| Thrum x Thrum | Ss x Ss | Incom. Mating | - |
TRISTYLY is known in some plants like Lythrum salicaria . In this case the style of the
flower may be short, long or medium length


Homomorphic System
Here the incompatibility is not associated with morphological difference among flower.
The incompatibility reaction of pollen may be controlled by the genotype of the plant on which it
is produced – (Sporophytic control) or by its own genotype – (Gametophytic control).
Gametophytic system
First discovered by East and Mangelsdorf in1925 in Nicotiana sanderae . Here the
incompatible reaction of pollen is determined by its own genotype and not by the genotype of the
plant on which pollen is produced. Genetically the incompatibility reaction is determined by a
single gene having multiple allele. Eg. Trifolium Nicotiana, Lycopersicon, Solanum,& Petunia .
Here codominance is assumed.
Genotype of plant S1S2 S3S4
(Sporophyte)
Genotype of gametes S1 S2 S3 S4
Incompatible
reaction of pollen S1 S2 S3 S4
Incompatible reaction
of style S1 S2 S3 S4
Mating S1S2 x S1S2 - Fully incompatible
S1S2 x S1S3 - Partially compatible
S1S2 x S3S4 - Fully compatible
Sporophytic system
Here also the self incompatibility is governed by a single gene S with multiple alleles.
More than 30 alleles are known in Brassica oleracea . Here dominance is assumed.
The incompatibility reaction isdetermined by the genotype of the plant on which pollen
grain is produced and not by the genotype of the pollen. This system is more complicated. The
allele may exhibit dominance, co-dominance or competition. This system was first reported by
Hugues and Babcock in1950 in Crepis foetida and by Gerstal in Parthenium argentatum . The
sporophytic system is found in radish, brassicas and spinach.
Lewis has summarized the characteristics of sporophytic system as follows :
-
There are frequent reciprocal differences
-
Incompatibility can occur with female parent
-
A family can consist of three incompatibility groups
-
Homozygotes are a normal part of the system
-
An incompatibility group may contain two genotypes.
Mechanism of Self Incompatibility
This is quite complex and is poorly understood. The various phenomena observed in Self
incompatibility is grouped into three categories.
-
Pollen – Stigma interaction
-
Pollen tube – Style interaction
-
Pollen tube – Ovule interaction
Pollen – Stigma interaction
This occurs just after the pollen grains reach the stigma and generally prevents pollen
from germination. Previously it was thought that binucleate condition of pollen in gametophytic
system and trinucleate condition in sporophyutic system was the reason for self incompatibility.
But later on it was observed that they are not the reason for SI. Under homomorphic system of
incompatibility there are differences in the stigmatic surface which prevents pollen germination.
In gametophytic system the stigma surface is plumose having elongated receptive cells which is
commonly known as wet stigma. The pollen grain germinates on reaching the stigma and
incompatibility reaction occurs at a later stage.
In the sporophytic system the stigma is papillate and dry and covered with hydrated layer
of protein known as pellicle. This pellicle is involved in incompatibility reaction. With in few
minutes of reaching the stigmatic surface the pollen releases an exine exudates which is either
protein or glycerol protein. This reacts with pellicle and induces callose formation, which further
prevents the growth of pollen tube.
| Gametophytic system | Sporophytic system |
|---|---|
| Stigma surface Plumose Commonly known as wet Stigma |
Stigma surface Papillate and dry. Covered with hydrated layers of protein known as pellicle which involves in incompatibility reaction |
| Pollen grain germinates and incompatibility reaction occurs at a later stage |
Pollen grain releases exine exudates which is protein or Glyco-protein |
| This protein reaction with pellicle and induces callose formation and arrests growth of pollen type. |
Pollen Tube – Style interaction
Pollen grains germinate and pollen tube penetrates the stigmatic surface. But in
incompatible combinations the growth of pollen tube is retarded with in the style as in Petunia,
Lycopersicon, & Lilium . The protein and poly saccharine synthesis in the pollen tube stops
resulting in bursting up of pollen tube and leading to death of nuclei.
Pollen tube – Ovule interaction
In Theobroma cacao pollen tube reaches the ovule and fertilization occurs but the
embryo degenerates later due to some biochemical reaction.
Male Sterility
Male sterility is characterized by nonfunctional pollen grains, while female gametes
function normally. It occurs in nature sporadically.
Morphological features of male sterility
The male sterility may be due to mutation, chromosomal aberrations, cytoplasmic factors or
interaction of cytoplasmic and genetic factors. Because of any of the above reasons the following
morphological changes may occur in male sterile plants.
- Viable pollen grains are not formed. The sterile pollen grains will be transparent and
rarely takes up stain faintly.
- Non-dehiscence of anthers, even though viable pollens are enclosed within. This may be
due to hard outer layer, which restrict the release of pollen grains.
-
Androecium may abort before the pollen grains are formed.
-
Androecium may be malformed, thus there is no possibility of pollen grain formation.
Kinds of male sterility, maintenance and uses
Male sterility may be conditioned due to cytoplasmic or genetic factors or due to
interaction of both. Environment also induces male sterility. Depending on these factors male
sterility can be classified in to
-
Cytoplasmic male sterility (CMS)
-
Cytoplasmic-genetic male sterility (CGMS)
-
Genetic male sterility (GMS)
In this there are two categories.
- Environment insensitive genic male sterility- commonly referred as Genetic male
sterility.
- Environment sensitive genic male sterility or Environmental induced sterility which is
again sub divided in to
-
TGMS (Thermosensitive)
-
PGMS (Photo sensitive)
-
Photo thermo sensitive
Cytoplasmic Male Sterility (CMS)
It occurs due to the mutation of mitochondria or some other cytoplasmic factors outside
the nucleus. Nuclear genes are not involved here. There is considerable evidence that gene or
genes conditioning cytoplasmic male sterility. Particularly in maize DNA reside in mitochondria
and may be located in a plasmid like element.
Genetic structure
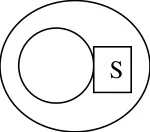
Sterile
Maintenance
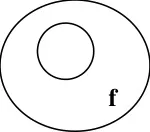
A line sterile B line fertile
Sterile
Since mother contributes the cytoplasm to the offspring, the sterility is transferred to the F1.
Uses
Since there are no R lines available, this type of sterility is useful only in crops where seed is
not the end product. For example in onion and many ornamental plants the hybrids developed
exhibit maximum hybrid vigour with respect to longer vegetative duration and larger flower size
and larger bulb size. Cytoplasmic male sterility has successfully been exploited in maize for
producing double cross hybrids.
A B C D
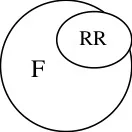
RR
Sterile (A x B) x (C x D)
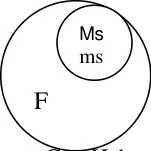
Genetic Male Sterility (Gms)
Genetic male sterility is normally governed by nuclear recessive genes ms ms. Exception to
this is safflower where male sterility is governed by dominant gene Ms Ms. This type of male
sterility is used in Redgram and Castor for production of hybrids.
Genetic structure
Maintenance
In genetic male sterility, the sterile line will be maintained from heterozygous condition.
sterile.
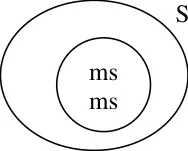
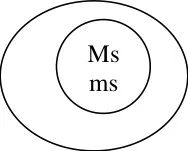
The pollen from the Fertile line will pollinate the sterile line and as a result seed set will
be there in the sterile line. These seeds are to be harvested and used for hybrid seed production.
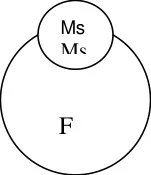
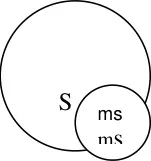
For hybrid seed production, the seeds collected from sterile plants will be grown using
double the seed rate since it will segregate in the ratio of 1 fertile : 1 sterile line. At the time of
flowering, the fertile line will be identified by yellow plumpy anthers and removed from the
field. Only the sterile line will remain in field. These will be pollinated by the R line and the R1
obtained will be hybrid redgram.
Utilisation: Hybrid development. Eg: Redgram
Ms T21 x ICPL 87109
A line R line
Difficulties in use of Gms
- Maintenance of GMS requires skilled labour to identify fertile and sterile line. Labelling
is time consuming and costly.
-
In hybrid seed production plot identification of fertile line and removing them is costly.
-
Use of double the seed rate of GMS line is costly
-
In crops like castor high temperature leads to break down of male sterility.
Transgenic Genetic Male Sterility
A gene introduced into the genome of an organism by recombinant DNA technology or
genetic engineering is called transgene. Many transgenes have been shown to produce genetic
male sterility, which is dominant to fertility. Consequently, it is essential to develop effective
fertility restoration system if these are to be utilized for hybrid seed production. An effective
restoration system is available in at least one case called Barnase or Barstar system.
The Barnase gene of Bacillus amyloliquefaciens encodes an RNAs. When Barnase gene is
driven by TA 29 promoter, it is expressed only in tapetum cells causing their degeneration.
Transgenic tobacco and Brassica napus plants expressing Barnase were completely male sterile.
Another gene, Barstar, from the same bacterium encodes a protein, which is a highly specific
inhibitor of Barnase RNAse. Therefore, transgenic plants expressing both Barstar and Barnase
are fully male fertile.
The Barnase gene has been tagged with bar gene, which specifies resistance to the herbaicde
phosphinothricin. This male sterile line is maintained by crossing with a male fertile line. The
progeny so obtained contain 1 male sterile : 1 male fertile plants; the latter are easily eliminated
at seedling stage by a phosphinothricin spray. The male sterile plants are crossed with the Barstar
line to obtain male fertile hybrid progeny. This system of male sterility is yet to be commercially
used.
Use of TGMS or PGMS eliminates this problem. These male sterile lines are maintained by
growing them in a locality where the temperatures and photoperiods during the sensitive
developmental stages are such that they exhibit complete male fertility (phenotypically).
The selfed seeds from such lines are then grown for hybrid seed production in those locations
where their ms genes produce complete male sterility due to the prevailing temperature and
photoperiod regimes. Clearly, all the plants in the male sterile line will be sterile and no rouging
will be required. TGMS and PGMS are being used for hybrid rice development in China.
Cytoplasmic Geneic Male Sterility
This is a case of cytoplasmic male sterility where dominant nuclear gene restorers fertility.
This system is utilized for the production of hybrids in bajra, jowar, maize, rice, wheat and many
other crops.
Genetic structure
A line
Male sterile
A line or ms line : This tem represents a male sterile line belonging to anyone of the above
categories. The A line is always used as a female parent in hybrid seed production.
B line or maintainer line : This line is used to maintain the sterility of A line. The B line is
isogenic line which is identical for all traits except for fertility status.
R line and restoration of fertility : It is other wise known as Restorer line which restores
fertility in the A line. The crossing between A x R lines results in F 1 fertile hybrid seeds which
is of commercial value.
Maintenance
A line B line
Sterile Fertile
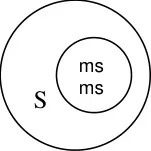
Male sterile line
The A line which is male sterile is maintained by crossing it with isogenic B line which is
also known as maintainer line. The B line is similar to that of A line in all characters
(isogenic) except fertile cytoplasm.
Utilsiation
The male sterile. A line is crossed with R line (Restorer) which restorers fertility in F1.
A line R line
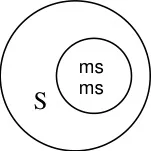
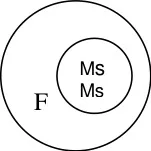
Sterile Fertile
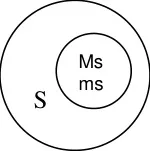
Hybrid Fertile
Limitations of CGMS lines
-
Fertility restoration is a problem. E.g. Rice.
-
Seed set will be low in crops like rice where special techniques are to be adopted to
increase seed set.
-
Break down of male sterility at higher temperature
-
In crops like wheat having a polyploidy series it is difficult to develop effective R line.
-
Undesirable effect of cytoplasm. Eg. Texas cytoplasm in maize became susceptible to
Helminthosporium. In bajra Tift 23 A cytoplasm became susceptible to downy mildew.
- Modifier genes may reduce effectiveness of cytoplasmic male sterility.
Summary Cheat Sheet
Quick Recall Points
- This lesson focuses on key plant breeding concepts, terminology, and exam-relevant applications.
- Review major definitions, classifications, and method-wise distinctions from the sections above.
- Revise tables and examples from this lesson for fast pre-exam recall.
Exam Traps
- Do not confuse similarly named breeding methods without checking their core selection logic.
- Pay attention to crop-specific examples because the same principle can behave differently by species.
References
1 source • [1]
References
Standard Plant Breeding Class Notes (GPBR211)
BookLesson Doubts
Ask questions, get expert answers