Understand the mechanism, types, and factors affecting crossing over, plus coincidence, interference, and significance in plant breeding — with agricultural examples and exam tips.
Mind Map: Crossing Over
Why Crossing Over Matters in Agriculture
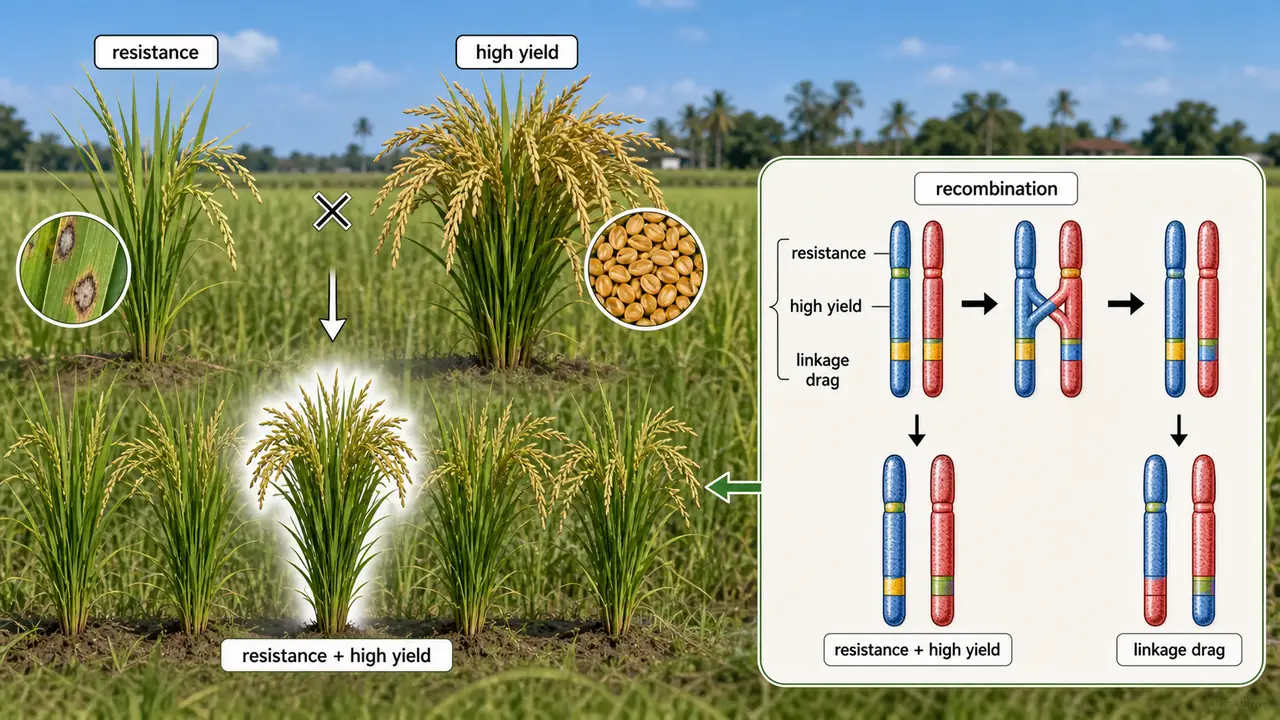
When a plant breeder crosses a blast-resistant rice variety with a high-yielding one, the desired outcome is offspring that carry both resistance and high yield. This is only possible because crossing over during meiosis breaks old gene combinations and creates new ones. Crossing over is also how breeders break linkage drag — separating a useful gene from an undesirable one that sits nearby on the same chromosome. Additionally, recombination frequencies from crossing over are used to construct genetic maps, which guide modern marker-assisted selection programmes.
Crossing over matters in breeding because it can create the exact recombinant plant that combines useful traits while shedding linkage drag.
What Is Crossing Over?
The term was first used by Morgan and Cattell (1912).
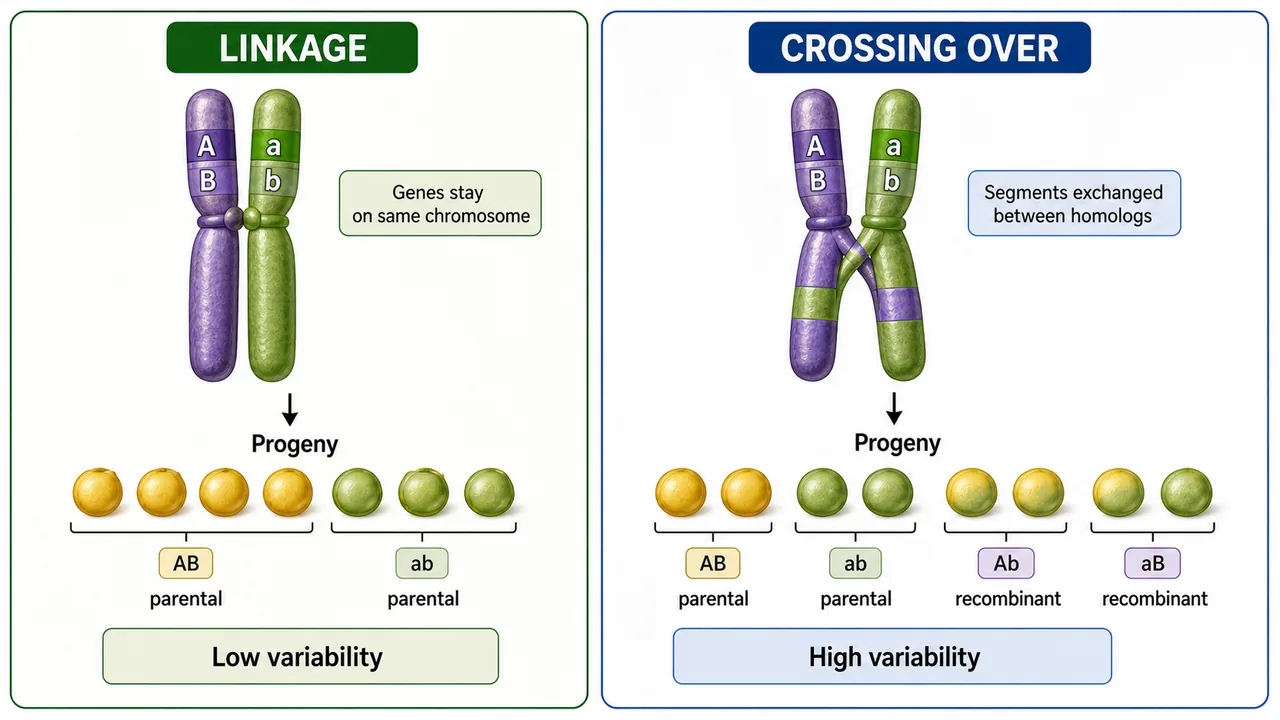
Crossing over is the physical exchange of precisely homologous segments between non-sister chromatids of homologous chromosomes during meiosis.
It produces recombinant (new) allele combinations that differ from the parental arrangements.
Crossing over — physical exchange of homologous segments between two non-sister chromatids at pachytene; produces recombinant chromatids with new allele combinations
Mechanism of Crossing Over
TIP
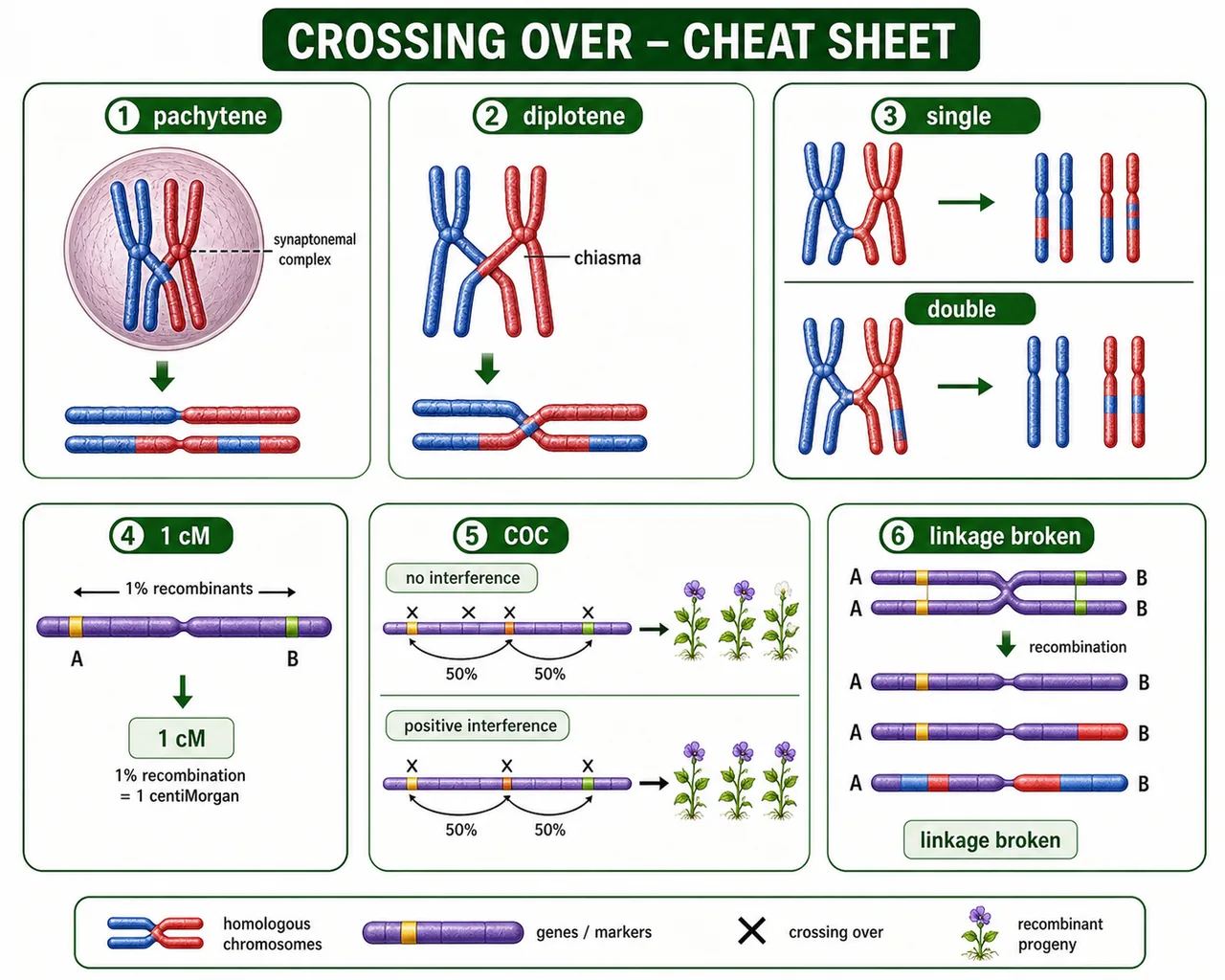
Crossing over occurs at pachytene, but chiasmata are visible at diplotene. This distinction is a common exam question.
Crossing over takes place during the pachytene stage of meiosis I — after homologous chromosomes have paired (synapsed) and before they begin to separate.
At pachytene, each bivalent has four chromatids (two per homologue) — called a tetrad or four-strand stage.
Breakage occurs at precisely homologous points in two non-sister chromatids, mediated by enzymes such as recombinase (endonuclease cuts, ligase seals).
The broken segments are exchanged and reunited — each crossing-over event involves two of the four chromatids.
This produces an X-shaped figure at the exchange point called a chiasma (plural: chiasmata), visible at diplotene.
Chiasma — the X-shaped figure formed at the site of crossing over; physical evidence of chromatid exchange; visible under the microscope at diplotene stage of meiosis I
Products of Crossing Over
Product
Description
Result
Crossover chromatids (2)
Participated in the exchange
Recombinant — new allele combinations
Non-crossover chromatids (2)
Did not participate
Parental — original allele combinations
Calculating Crossing Over Frequency
Crossing over frequency formula — recombination % = (recombinant progeny ÷ total progeny) × 100; 1% recombination = 1 centiMorgan (1 map unit)
Crossing over % = (Recombinant progeny / Total progeny) x 100
The recombination frequency is a direct measure of genetic distance between two genes: 1% recombination = 1 map unit (centiMorgan).
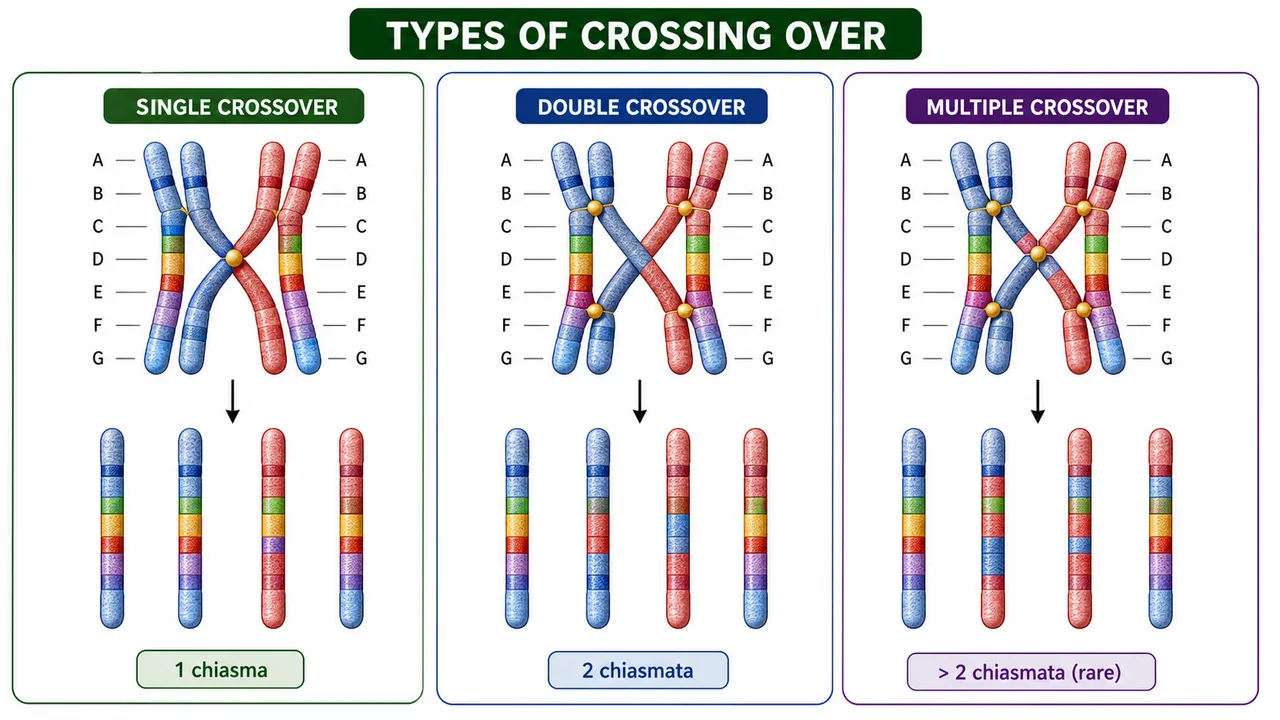
Types of Crossing Over
Type
Chiasmata
Genes Involved
Test Cross Type
Frequency
Single
1 chiasma
2 linked genes
Two-point test cross
Most common
Double
2 chiasmata
3 linked genes
Three-point test cross
Less common; used to determine gene order
Multiple
>2 chiasmata
>3 linked genes
—
Extremely rare (due to interference)
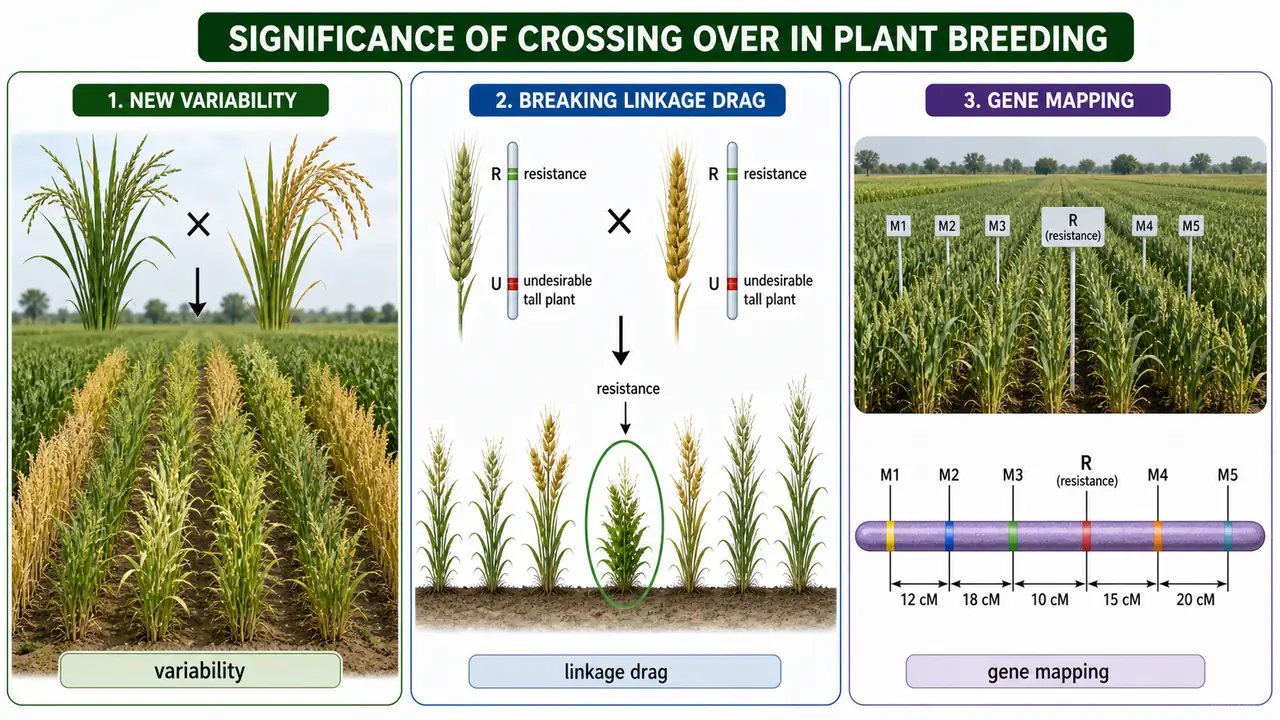
Agricultural application: Three-point test crosses are used in genetic mapping to determine gene order and calculate map distances between three genes simultaneously — essential for constructing linkage maps in crop species.
This board separates single, double, and multiple crossovers by the number of chiasmata and the recombinant products they can generate.
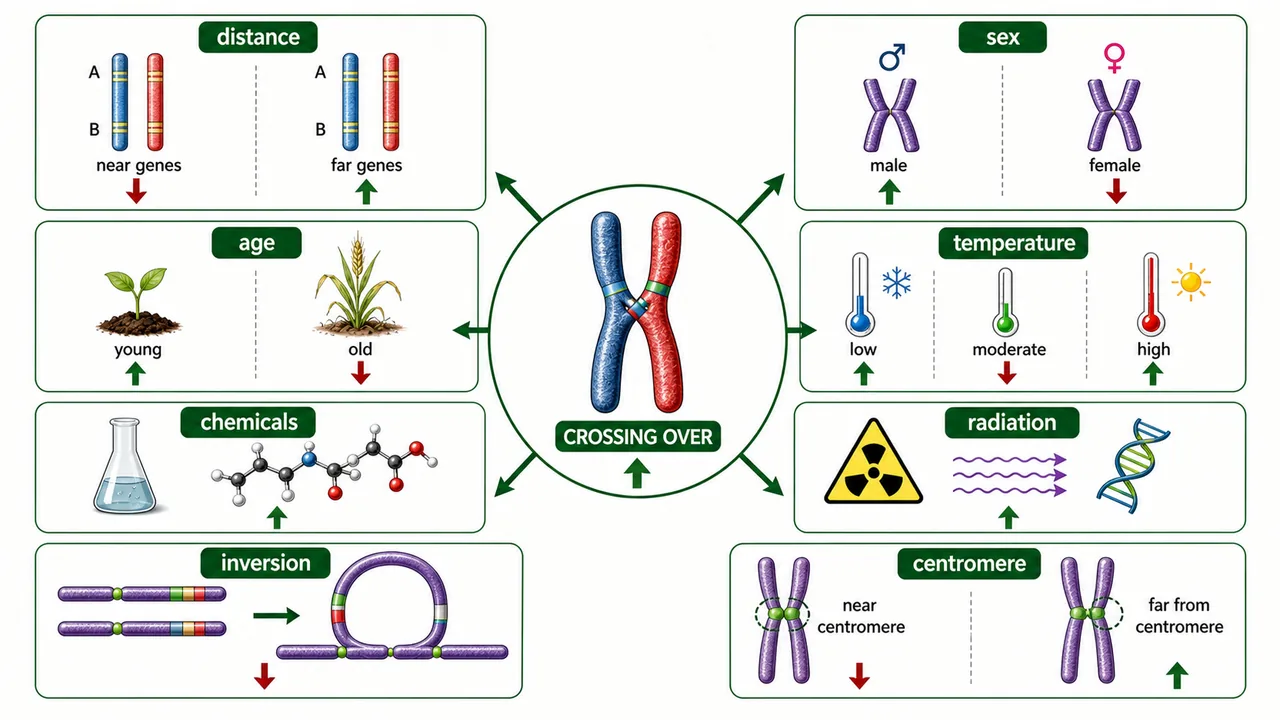
Factors Affecting Crossing Over
Factor
Effect on Crossing Over
Mechanism
Distance between genes
Increases with distance (positively correlated)
Basis of genetic mapping: 1% CO = 1 cM
Sex
Heterogametic sex shows lower CO
No CO in Drosophila males or silkworm females
Age of female
Declines with advancing age
Recombination machinery becomes less efficient
Temperature
Lowest at 22°C in Drosophila; increases at extremes
Temperature stress disrupts normal CO control
Nutrition (Ca²⁺, Mg²⁺)
Higher metallic ions → lower CO
Divalent cations affect chromosome structure
Chemicals
Mitomycin D, actinomycin D, EMS → increase CO
Interfere with DNA replication and repair
Radiation
X-rays, gamma rays → increase CO
Radiation-induced DNA breaks trigger recombination
Plasmagenes
Some cytoplasmic genes reduce CO
E.g., Tifton male-sterile cytoplasm reduces CO in bajra
Genotype
Some genes promote or inhibit CO
C3G gene in Drosophila: homozygous = prevents CO; heterozygous = promotes CO
Chromosomal aberrations
Inversions reduce CO within inverted segment
Recombinant products from inversions are often inviable
Crossing-over frequency changes with chromosome context, genotype, and environment, so breeders treat recombination as a controllable tendency rather than a fixed constant.
Significance in Plant Breeding
For plant breeding, crossing over is valuable because it creates new variation, breaks undesirable linkage, and supports gene mapping.
Significance
Detail
Increases variability
Creates new allele combinations — the genetic diversity breeders select from
Breaks linkage
Separates desirable genes from undesirable ones (linkage drag)
Agricultural example: In wheat, crossing over has been used to break the linkage between a disease-resistance gene from a wild relative (Aegilops) and genes for poor grain quality — a classic case of overcoming linkage drag through recombination.
Coincidence and Interference
Coincidence
Refers to the occurrence of two or more crossovers simultaneously in the same chromosomal region → double crossover product.