🧬 Somatic Embryogenesis
Meaning, features, and crop improvement relevance of somatic embryogenesis.
Somatic embryogenesis is a key tissue culture concept because it proves that a somatic cell can be redirected to form an embryo-like structure capable of regenerating into a whole plant. This makes it important for regeneration systems, mass propagation, and synthetic seed work.
Somatic embryogenesis
The sporophytic generation of a plant is initiated with the zygote, which is the initial cell
(product of gamete fusion) that bears all the genetic information to construct the adult
individual. In angiosperms, the zygote divides transversally, resulting in two cells. One of
them, the apical cell, is small and dense with an intense activity of DNA synthesis. Further
ordinal divisions of this cell give rise to the embryo head that will be the new plant. The
other resultant cell (basal cell) is a large and highly vacuolated one that will confirm the
suspensor complex, which plays an important role during the early stages of the young
embryo. In vivo -produced somatic embryos generally follow the same pattern; however,
they are not initiated from a zygote, but from a somatic cell as occurs in the polyembryony.
Somatic embryos
Somatic embryos are bipolar structures with both apical and basal meristematic regions,
which are capable of forming shoot and root, respectively. A plant derived from a somatic
embryo is sometimes referred to as an “embling”.
Somatic embryos vs zygotic embryos and their advantages
Somatic embryos are structurally similar to zygotic embryos found in seeds and possess
many of their useful features, including the ability to grow into complete plants. However,
somatic embryos differ in that they develop from somatic cells, instead of zygotes (i.e.,
fusion product of male and female gametes) and thus, potentially can be used to produce
duplicates of a single genotype. Since the natural seed develops as a result of a sexual
process in cross-pollinating species, it is not genetically identical to one single parent. In
contrast, somatic embryo develops from somatic cells (non-sexual) and does not involve
sexual recombination. This characteristic of somatic embryos allows not only clonal
propagation but also specific and directed changes to be introduced into desirable elite
individuals by inserting isolated gene sequences into somatic cells. This bypasses
genetic recombination and selection inherent in conventional breeding technology. If the
production efficiency and convenience comparable to that of a true seed are achieved,
somatic embryos can be potentially used as a clonal propagation system.
Somatic embryogenesis can be initiated by two mechanisms: directly on explanted
tissues, where plants are genetically identical (clonation), and indirectly from unorganized
tissues (callus). Propagation by indirect embryogenesis carries the risk of producing
plants that may differ genetically from each other and from the parental plant. It is thought
that the occurrence of genetic variability within tissue cultures may partly originate from
cellular changes that are induced during culture. The genetic variability associated with
tissue and cell culture is named "somaclonal variation" and represents an opportunity
where selection pressure can be imposed to isolate unique genetic forms of a clone. The
ability to recover plants from single cells has made possible the genetic improvement in
vitro . Plant tolerance to abnormal temperatures, herbicides, fungal toxins, high levels of
salt, etc., can be obtained by exposing cell cultures to a selective agent. The most
important advantages of the cell- and tissue-culture tools used in plant breeding are:
-
Freedom from the effects of the natural environment.
-
The ability to handle large numbers of individuals (cells) in very small spaces.
-
Creation of variation within cultures. Variability usually occurs spontaneously; however, it
can be induced by mutagenic agents.
- Genetic variability can be created deliberately in cultured cells by using
genetic-engineering techniques.
Practical applications of somatic embryogenesis
Clonal propagation
Somatic embryogenesis has a potential application in plant improvement. Since both the
growth of embryogenic cells and subsequent development of somatic embryos can be
carried out in a liquid medium, it is possible to combine somatic embryo-genesis with
engineering technology to create large-scale mechanised or automated culture systems.
Such systems are capable of producing propagules (somatic embryos) repetitively with
low labour inputs. In this process of repetitive somatic embryogenesis (also referred to as
accessory, adventive, or secondary somatic embryogenesis) a cycle is initiated whereby
somatic embryos proliferate from the previously existing somatic embryo in order to
produce clones.
Cloning zygotic embryos for repetitive somatic embryogenesis
A wide range of soybean genotypes have been tested for their ability to undergo
auxin-stimulated somatic embryogenesis during cloning of zygotic embryos (Barwale et
al . 1986; Komatsuda and Ohyama 1988). All of them are reported to form somatic
embryos provided appropriate nutrients are provided in the medium. The role of
genotypes in conferring regeneration capacity is further supported by studies on zygotic
embryo cloning of wheat, rice and maize. Analysis of various cultivars demonstrated that
the regeneration capacity of these crops was directly affected by non-additive, additive
and cytoplasmic factors. However, the genotype which has the capacity to undergo
repetitive somatic embryogenesis can be back-crossed to elite lines in order to transform
the latter with capacity for high regeneration of somatic embryos. Such a type of
transformation could play an important role in plant breeding since through in vitro
techniques high quality somatic embryos have been produced in 80 species of tropical
crops.
Raising somaclonal variants in tree species
Embryos formed directly from pre embryogenic cells appear to produce relatively uniform
clonal material, whereas the indirect pathway generates a high frequency of somaclonal
variants. Mutation during adventive embryogenesis may give rise to a mutant embryo
which on germination would form a new strain of plant. Nucellar embryos, like shoot tips,
are free of virus and can be used for raising virus-free clones, especially from some tree
species (e.g., poly embryonate Citrus) where shoot tip culture has not been successful.
For clonal propagation of tree species, somatic embryogenesis from nucellar cells may
offer the only rapid means of obtaining juvenile plants equivalent to seedlings with
parental genotype. Clonal propagation through somatic embryogenesis has been
reported in 60 species of woody trees representing 25 families. Somatic embryos with
genotype of a selected elite parent are potentially convenient organs for cryopreservation
and germplasm storage.
Synthesis of artificial seeds
There development of methods for encapsulation of somatic embryos to enable them to
be sown under field conditions as „synthetic‟ or „artificial‟ seed is gaining importance.
Research programmes on production of artificial seeds via somatic embryogenesis in
respect of commercially important crops would not only contribute to increased
agricultural production, but also add to our basic knowledge of the regulatory mechanisms
which control plant growth and differentiation.
Source of regenerable protoplast system
Embryogenic callus, suspension cultures and somatic embryos have been employed as
sources of protoplast isolation for a range of species. Cells or tissues in these systems
have demonstrated the potentiality to regenerate in cultures and, therefore, yield
protoplasts that are capable of forming whole plants. Embryogenic cultures are especially
valuable in providing a source of regenerable protoplasts in the graminaceous, coniferous
and citrus species. Attempts to achieve regeneration of callus or even sustained divisions
in mesophyll-derived protoplasts of Gramineae proved unsuccessful until Vasil and Vasil
(1980) turned to embryogenic cultures obtained from immature embryos of pearl millet
( Pennisetum purpureum ) as the source of protoplasts. Protoplasts from these cultures
were induced to divide to form a cell mass from which embryoids, and even plantlets,
regenerated on a suitable nutrient medium. Similar success was subsequently reported by
other workers with embryogenic suspension of Panicum maximum, Pennisetum
purpureum, Oryza sativa, Saccharum officinarum, Lolium perenne, Festuca arundinaceae
and Dactylis glomerata . Among cereals, in Gramineae, the development of a protoplast
regeneration system for maize has been especially challenging. Rhodes et al . (1988) were
the first to raise protoplast derived maize plantlets from embryogenic cultures which,
however, proved sterile. Later, Shillito et al . (1989) initiated embryogenic callus cultures
from immature zygotic embryos of an elite inbred maize line B73, which yielded
protoplasts that regenerated to form fertile plants. The protoplast-derived plant on
crossing with pollen from a seed-derived plant produced viable seeds which germinated
normally. Using similar techniques, Prioli and Sondahl (1989) obtained fertile maize plants
from embryogenic cell suspension protoplasts of a line (Cat 100-1) adapted to tropical
regions.
Embryogenic citrus suspension cultures also provide protoplasts that can be used in the
production of interspecific and intergeneric somatic hybrid plants. Equal success can be
obtained with protoplasts isolated from nucellus-callus culture of this plant. Ability to
isolate protoplasts from embryogenic cultures has had a large impact on their in vitro
culture of forest trees, e.g., Pinus taeda, Picea glauca, P. mariana, Pseudotsuga
menziesii, Abies alba, Santalum album and Liriodendron tulipfera . Somatic embryos
induced on the protoplast-derived calli also germinate to form plantlets which finally
establish in the soil.
Genetic transformation
In seed embryogenesis, zygotic embryos are seated deep inside the nucellar tissue. They
live in a protected environment besides being genetically heterogeneous. On the contrary,
somatic embryos remain virtually unprotected and more or less give rise to genetically
uniform plants. The advent of leaf-disc transformation systems has made it possible to
successfully engineer species ( Nicotiana tabacum, Medicago sativa ) in which, tissues are
capable of regeneration via somatic embryogenesis.
In these species, isolated single cells can be transformed in cultures and grown on a
selection medium (nutrient medium containing an antibiotic, kanamycin) to callus colonies
which eventually form somatic embryos on removal of auxin from this medium (Chabaud
et al ., 1988). Since the callus phase seems essential in this type of indirect somatic
embryogenesis, the possibility of chimeric embryos arising from transformed and
non-transformed tissues cannot be ruled out. Therefore, the callus phase can be
bypassed through a process of repetitive somatic embryogenesis. McGranahan et al .
(1990) used repetitive embryogenesis for Agrobacterium -transformed walnut ( Juglans
regia L.) cells and obtained multiple crops of somatic embryos without employing the
callus phase.
There is also evidence to show that repetitive embryos originate from single epidermal or
subepidermal cells which can be readily exposed to Agrobacterium . Thus, the
transformation technique applied to a primary somatic embryo, instead of a zygotic
embryo, should give rise to totally transgenic somatic embryos. Repetitive embryogenesis
is also ideally suited to particle gun-mediated genetic transformation. Instead of relying on
Agrobacterium to mediate the transfer of genes into plant cells, the particle gun literally
shoots DNA that has been precipitated onto particles of a heavy metal, into the plant cells.
Embryogenic suspension cultures of cotton and soybean, initiated from immature
embryos, yielded an average of 30 stably transformed cell lines following each firing of the
gun. The transformed cell lines can then be induced to form an unlimited number of
transformed somatic embryos through repetitive embryogenesis.
Synthesis of metabolites
The repetitive embryogenesis system is of potential use in the synthesis of metabolites
such as pharmaceuticals and oils. Borage ( Borage officinalis L.) seeds contain high levels
of ylinolenic acid, used in the treatment of atopic eczema. Somatic embryos of borage also
produce this metabolite but through repetitive somatic embryogenesis a continuous
supply of y-linolenic acid is ensured, which otherwise would be limited to the growing
season in the zygotic embryo. The same principle can be applied for production in vitro of
industrial lubricants from jojoba ( Simmondsia chinensis ) and leo-palmitostearin (the major
ingredient in cocoa butter) from Cacao ( Theobroma cacao ).
The in vivo clonal propagation of plants is often difficult, expensive and even
unsuccessful. Tissue culture methods offer an alternative means of plant vegetative
propagation. Clonal propagation through tissue culture (popularly called
micropropagation) can be achieved in a short time and space. This is possible through
somatic embryogenesis by which rapid formation of embryos takes place leading to
multiplication of shoots.
Synthetic seed production
Need for artificial or synthetic seed production technology
Development of micropropagation techniques will ensure abundant supply of the desired
plant species. In some crop species seed propagation has not been successful. This is
mainly due to heterozygosity of seed, minute seed size, presence of reduced endosperm
and the requirement of seed with mycorrhizal fungi association for germination (eg.
orchids), and also in some seedless varieties of crop plants like grapes, watermelon, etc.
Some of these species can be propagated by vegetative means. However, in vivo
vegetative propagation techniques are time consuming and expensive. Development of
 artificial seed production technology is currently considered as an effective and efficient
artificial seed production technology is currently considered as an effective and efficient
alternate method of propagation in several commercially important agronomic and
horticultural crops. It has been suggested as a powerful tool for mass propagation of elite
plant species with high commercial value. Characteristics of clonal propagation systems
are discussed in Box 1.
Artificial seed technology involves the production of tissue culture derived somatic
embryos encased in a protective coating. Artificial seeds have also been often referred to
as synthetic seeds. However, the term synthetic seed should not be confused with
commercial seeds of a synthetic cultivar which is defined as an advanced generation of an
open pollinated population composed of a group of selected inbred clones or hybrids. The
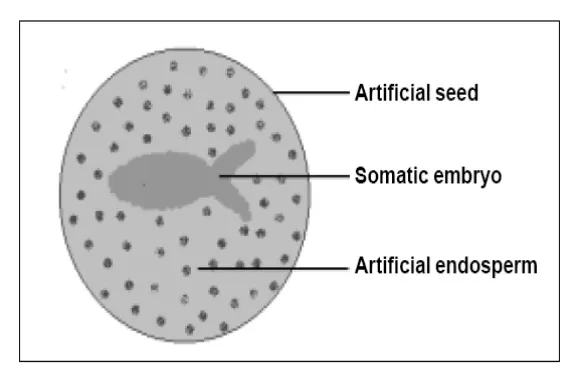
concept of artificial or synthetic seed is shown in Figure.
These synthetic seeds would
also be a channel for new plant
lines produced through
biotechnological advances to
be delivered directly to the
greenhouse or field.
Advantages of artificial/
synthetic seeds over somatic
embryos for propagation are
listed in Box 2. This synthetic
seed production technology is a high volume, low-cost production technology. High
volume propagation potential of somatic embryos combined with formation of synthetic
seeds for low-cost delivery would open new vistas for clonal propagation in several
commercially important crop species.

Basic requirement for production of artificial seeds
Recently, production of synthetic seeds by encapsulating somatic embryos has been
reported in few species. One prerequisite for the application of synthetic seed technology
in micropropagation is the production of high-quality, vigorous somatic embryos that can
produce plants with frequencies comparable to natural seeds. Inability to recover such
embryos is often a major limitation in the development of synthetic seeds.
Synthetic seed technology requires the inexpensive production of large numbers of high
quality somatic embryos with synchronous maturation. The overall quality of the somatic
embryos is critical for achieving high conversion frequencies. Encapsulation and coating
systems, though important for delivery of somatic embryos, are not the limiting factors for
development of synthetic seeds.
At present, the characteristic lack of developmental synchrony in embryogenic systems
stymies multi-step procedures for guiding somatic embryos through maturation. The lack
of synchrony of somatic embryos is, arguably, the single most important hurdle to be
overcome before advances leading to widespread commercialization of synthetic seeds
can occur. Synchronized embryoid development is required for the efficient production of
synthetic seeds.
Procedure for production of artificial seeds
Establish somatic embryogenesis
↓
Mature somatic embryos
↓
Synchronize and singulate somatic embryos
↓
Mass production of somatic embryos
↓
Standardization of encapsulation
↓
Standardization of artificial endosperm
↓
Mass production of synthetic seeds
↓
Greenhouse and field planting
Types of somatic embryos
Two types of artificial seeds have been developed, namely, hydrated and desiccated.
Redenbergh et al. (1986) developed hydrated artificial seeds by mixing somatic embryos
of alfalfa, celery and cauliflower with sodium alginate, followed by dropping into a solution
of calcium chloride to form calcium-alginate beads. The beads become hardened as
calcium alginate is formed. After about 20-30 min. the artificial seeds are removed,
washed with water and used for planting. Hydrated artificial seeds are sticky and difficult to
handle on a large scale, dry rapidly in the open air. These problems can be resolved by
providing a waxy coating over the beads. However, it is not possible to store, except at low
temperatures and for limited periods, hydrated artificial seeds and they have to be planted
soon after they are produced. Precision machines for large scale encapsulation of SEs
have been devised.
Kim and Janick (1989) applied synthetic seed coats to clumps of carrot somatic embryos
to develop desiccated artificial seeds. They mixed equal volumes of embryo suspension
and 5% solution of polyethylene oxide (polyox WSR N-750), a water-soluble resin, which
subsequently dried to form polyembryonic desiccated wafers. The survival of
encapsulated embryos was further achieved by embryo hardening treatments with 12%
sucrose or10-6M ABA, followed by chilling at high inoculum density.
Another delivery system for somatic embryos for obtaining transgenic plants is Fluid
drilling. Embryos are suspended in a viscous-carrier gel which extrudes into the soil. The
primary problem in fluid-drilling is that the sucrose level necessary to permit conversion
also promotes rapid growth of contaminating micro-organisms in a non-aseptic system.
Gray (1987) found that somatic embryos of orchard grass ( Dactylis glomerata ) became
quiescent when desiccated in empty plastic petri dishes at 70% relative humidity at 23°C
which amounted to loss of 13% water. However, after 21 days of storage, desiccated
embryos when rehydrated in vitro germinated to produce viable plantlets though limited
(4%) in number. Senaratna et al (1990) treated alfalfa somatic embryos with ABA at the
torpedo to cotyledonary stages in order to increase their tolerance to desiccation. Over
60% of such desiccated embryos germinated when placed on a moist filter paper or sown
directly onto sterile soil and formed plantlets. Heat-shock treatments also induced a
degree of desiccation tolerance comparable to that conferred by ABA treatment and had
no detrimental effect on the subsequent growth of the plantlets.
In general, the gels contain inorganic nutrients, a carbon source, fungicides, bactericides
and other growth promoting substances necessary for SE germination and seedling
establishment in the field. When shoot buds are encapsulated, growth regulators needed
for rooting and shoot growth are also included. The SE must be of good quality to be able
to germinate in a high frequency under field/ greenhouse conditions. Alfalfa is the most
extensively investigated plant species in this regard.
Types of gelling agents used for encapsulation
Several gels like agar, alginate, polyco 2133 (Bordon Co.), carboxy methyl cellulose,
carrageenan, gelrite (Kelko. Co.), guargum, sodium pectate, tragacanth gum, etc. were
tested for synthetic seed production, out of which alginate encapsulation was found to be
more suitable and practicable for synthetic seed production. Alginate hydrogel is
frequently selected as a matrix for synthetic seed because of its moderate viscosity and
low spin ability of solution, low toxicity for somatic embryos and quick gelation, low cost
and bio-compatibility characteristics. The use of agar as gel matrix was deliberately
avoided as it is considered inferior to alginate with respect to long term storage. Alginate
was chosen because it enhances capsule formation and also the rigidity of alginate beads
provides better protection (than agar) to the encased somatic embryos against
mechanical injury. Alginate encapsulated somatic embryos of orchids are shown in Figure
1 and the plantlets derived from artificial or synthetic seeds of orchid are shown in Figure
Figure 1 (left). Artificial or synthetic seed produced in orchids by alginate encapsulation.
Figure 2 (right). Artificial or synthetic seed derived plantlets in orchid.
Principle and conditions for encapsulation with alginate matrix
Alginate is a straight chain, hydrophilic, colloidal polyuronic acid composed primarily of
hydro--D-mannuronic acid residues with 1-4 linkages. The major principle involved in
the alginate encapsulation process is that the sodium alginate droplets containing the

 somatic embryos when dropped into the CaCl2.2H2O solution form round and firm beads
somatic embryos when dropped into the CaCl2.2H2O solution form round and firm beads
due to ion exchange between the Na+ in sodium alginate with Ca [2+] in the CaCl2.2H2O
solution. The hardness or rigidity of the capsule mainly depends upon the number of
sodium ions exchanged with calcium ions. Hence, the concentration of the two gelling
agents i.e., sodium alginate and CaCl2.2H2O, and the complexing time should be
optimized for the formation of the capsule with optimum bead hardness and rigidity. In
general, 3% sodium alginate upon complexation with 75 mM CaCl2.2H2O for half an hour
gives optimum bead hardness and rigidity for the production of viable synthetic seeds.
Artificial endosperm
Somatic embryos lack seed coat (testa) and endosperm that provide protection and
nutrition for zygotic embryos in developing seeds. To augment these deficiencies,
addition of nutrients and growth regulators to the encapsulation matrix is desired, which
serves as an artificial endosperm. Addition of nutrients and growth regulators to the
encapsulation matrix results in increase in efficiency of germination and viability of
encapsulated somatic embryos. These synthetic seeds can be stored for a longer period
of time even up to 6 months without losing viability, especially when stored at 4 [0] C.
Addition of adjuvants to the matrix
In addition to preventing the embryo from desiccation and mechanical injury, a number of
useful materials such as nutrients, fungicides, pesticides, antibiotics and microorganisms
(eg. rhizobia) may be incorporated into the encapsulation matrix. Incorporation of
activated charcoal improves the conversion and vigour of the encapsulated somatic
embryos. It has been suggested that charcoal breaks up the alginate and thus increases
respiration of somatic embryos (which otherwise lose vigour within a short period of
storage). In addition, charcoal retains nutrients within the hydrogel capsule and slowly
releases them to the growing embryo.
Utilization of artificial seeds
The artificial seeds can be used for specific purposes, notably multiplication of non-seed
producing plants, ornamental hybrids (currently propagated by cuttings) or the
propagation of polyploid plants with elite traits. The artificial seed system can also be
employed in the propagation of male or female sterile plants for hybrid seed production.
Cryo-preserved artificial seeds may also be used for germplasm preservation, particularly
in recalcitrant species (such as mango, cocoa and coconut), as these seeds will not
undergo desiccation. Furthermore, transgenic plants, which require separate growth
facilities to maintain original genotypes may also be preserved using somatic embryos.
Somatic embryogenesis is a potential tool in the genetic engineering of plants. Potentially,
a single gene can be inserted into a somatic cell. In plants that are regenerated by somatic
embryos from a single transgenic cell, the progeny will not be chimeric. Multiplication of
elite plants selected in plant breeding programs via somatic embryos avoids the genetic
recombination, and therefore does not warrant continued selection inherent in
conventional plant breeding, saving considerable amount of time and other resources.
Artificial seeds produced in tissue culture are free of pathogens. Thus, another advantage
is the transport of pathogen free propagules across the international borders avoiding bulk
transportation of plants, quarantine and spread of diseases.
Potential uses of artificial seeds
Delivery systems
-
Reduced costs of transplants
-
Direct greenhouse and field delivery of:
-
Elite, select genotypes
-
Hand-pollinated hybrids
-
Genetically engineered plants
-
Sterile and unstable genotypes
-
-
Large-scale mono cultures
-
Mixed-genotype plantations
-
Carrier for adjuvants such as microorganisms, plant growth regulators pesticides,
fungicides, nutrients and antibiotics
-
Protection of meiotically-unstable, elite genotypes
-
Can be conceivably handled as seed using conventional planting equipment.
Analytical tools
-
Comparative aid for zygotic embryogeny
-
Production of large numbers of identical embryos
-
Determination of role of endosperm in embryo development and germination
-
Study of seed coat formation
-
Study of somaclonal variation.
Applicability and feasibility of artificial seed production technology
In order to be useful, synthetic seed must either reduce production costs or increase crop
value. The relative benefits gained, when weighed against development costs, will
determine whether its use is justified for a given crop species. Considering a combination
of factors, including improvement of the existing embryogenic systems, relative cost of
seed as well as specific application for synthetic seed allows judgement of relative need
for a given crop. For example, synthetic seed of seedless watermelon would actually cost
less than conventional seed, providing a benefit at the outset of crop production. Although
embryogenic systems for this crop do not exist, the benefit that could be conferred by use
of synthetic seed would be very great. Value added aspects that would increase crop
worth are numerous and include cloning of elite genotypes, such as genetically
engineered varieties, that cannot produce true seed.
Questions
- A plant derived from a somatic embryo is referred as …………
a). Seedling b). Embling c). Both a and b d). None of the above
- Somatic embryos are …………
a). Bipolar structures b).Have apical and basal meristematic regions c). Capable of forming shoot and d). All the above root
- Synthetic seeds are of ………… types
a). Hydrated b).Dessicated c). Both a and b d). None of the above
- Hydrated synthetic seeds are developed by ………..
a). Redenbergh b). Kim and Janick c). Both a and b d). None of the above
- Dessicated synthetic seeds are developed by ………..
a). Redenbergh b). Kim and Janick c). Both a and b d). None of the above
- Hydrated artificial seeds are ………..
a). Sticky b). Dry rapidly in the open air c). Difficult to handle on a large scale d). All the above
- Gels used for synthetic seed production includes….……..
a). Agar and alginate b). Polyco 2133 c). Carboxy methyl cellulose d). All the above
- Gels used for synthetic seed production includes….……..
a). Carrageenan and gelrite b). Sodium pectate c). Guargum d). All the above
- Alginate hydrogel is frequently selected as a matrix for synthetic seed because of its
….……..
a). moderate viscosity and low spin ability of solution
a). moderate viscosity and low b). low toxicity for somatic spin ability of solution embryos and quick gelation
c). low cost and bio-compatibility d). All the above characteristics
d). All the above
- Gels used for synthetic seed production includes….……..
a). Carrageenan and gelrite b). Sodium pectate c). Guargum d). All the above
- Incorporation of activated charcoal in synthetic seed coating improves ……..
a). Conversion and vigour of the encapsulated somatic embryos b). Increases respiration of somatic embryos c). Retains nutrients within the hydrogel capsule and slowly releases them to the growing embryo d). All the above
- The artificial seeds can be used for specific purposes viz., ……..
a). Multiplication of non-seed producing plants b). Propagation of male or female sterile plants for hybrid seed production c). Propagation of polyploid plants with elite traits d). All the above
Summary Cheat Sheet
Quick Recall Points
- Define key terms in one line and revise their use in plant biotechnology.
- Memorize major steps, methods, and applications covered in this lesson.
- Practice exam-style distinctions between related concepts and techniques.
Exam Traps
- Do not confuse similar terms without checking context and biological level.
- Revise process order carefully; sequence-based questions are common.
- Link each method with its most likely application question.
References
1 source • [1]
References
Standard BSc Agriculture Plant Biotechnology notes
BookLesson Doubts
Ask questions, get expert answers