🧬 Protoplast Isolation and Fusion
Isolation of plant protoplasts, fusion methods, and their significance in somatic hybridization.
This lesson explains why protoplasts matter in biotechnology. Once the rigid cell wall is removed, the plant cell becomes accessible for fusion, DNA uptake, and somatic hybridization, which is especially useful when normal crossing is difficult or impossible.
Why Protoplasts Matter
In eukaryotes the transfer of genetic material form one individual to another is conventionally
achieved through sexual breeding. In plants, where fairly distant species could be crossed, it
has not always been possible to obtain full hybrids between desired individuals because of
sexual incompatibility barriers. In this respect cell fusion offers a novel approach to distant
hybridization through somatic hybridization. Fusion of cells must occur through the plasma
membrane. Unlike animals, in plants the plasma membrane is bound by a rigid cellulosic wall
and the adjacent cells are cemented together by a pectin rich matrix. That’s why somatic cell
genetics is more advanced in animals than plants. In 1960, E. C. Cocking demonstrated the
feasibility of enzymatic degradation of plant cell walls to obtain large quantities of viable naked
cells – called as protoplasts.
Besides being able to fuse with each other, higher plant protoplasts can also take up foreign
DNA, through their naked plasma membrane under specific chemical and physical treatments.
Protoplasts also provide an experimental system for a wide range of biochemical and molecular
studies ranging from investigations into the growth properties of individual cells to membrane
transport.
Isolation of Protoplasts
Mechanical method
Klecker in 1892 has first initiated the protoplast isolation by mechanical means- the cells
were kept in a suitable medium plasmolyticum and cut with a fine knife. In this process some of
the plasmolyzed cells were cut only through the cell wall, releasing intact protoplasts.
Principle of Mechanical Method Isolation when tissue is cut along the dotted line, with a
razor blade, some protoplasts will be released
A. Tissue is cut along the dotted line B. Release of Protoplasts from Damaged cells
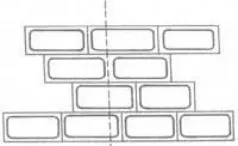
Limitation
-
Applicable only to vacuolated cells
-
Yields are extremely low.
Enzymatic method
In 1960, Cocking used a concentrated solution of cellulase enzyme, prepared from cultures of
the fungus, Myrothecium verrucaria, to degrade the cell walls.
However, real progress in this area was made after 1968 when cellulase and macerozyme
enzymes became commercially available. The commercial preparations of the enzymes for
protoplast isolation were first employed by Takebe et al ., (1968). The tobacco leaf species were
first exposed to macerozyme to liberate single cells which were then treated with cellulase to
digest the cell walls and release the protoplasts. Later, these two enzymes were used together
and this is found as faster method and also reduces the chances of microbial contamination by
cutting down a few steps.
Isolation of Mesophyll Protoplasts Using Leaves with Epidermis Peeled Off
- Leaves, 2. Sterilization of leaves, 3. Removal of epidermis by peeling, 4. Peeled leaves in
enzyme solution, 5. Released protoplasts in enzyme solution, 6. Protoplasts in washing
medium, 7. Centrifugation, 8. Cell debris, 9. Protoplasts and 10. Isolated protoplasts
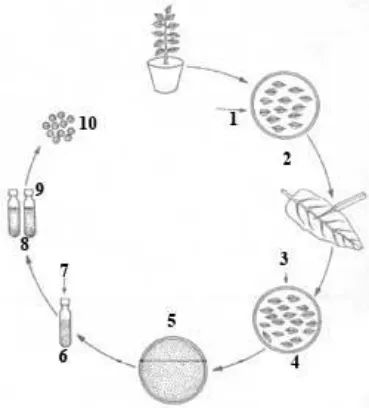 Table: Some commonly used commercially available enzymes for protoplast isolation
Table: Some commonly used commercially available enzymes for protoplast isolation
| Enzymes | Source |
|---|---|
| Cellulases Onozuka RS Cellulase R-10 |
Trichoderma viride T. viride |
| Hemicellulase Hemicellulase Rhozyme HP150 |
Aspergillus niger A. niger |
| Pectinase Macerozyme R-10 Macerase |
Rhizopus spp. Rhizopus spp. |
A range of enzyme preparations are now available commercially and depending on the nature
of the tissue these are used in different combinations. The use of commercially available
enzymes has enabled the isolation of protoplasts from virtually every plant tissue as long as
cells have not acquired lignification. Protoplast isolation has been reported from mesophyll cells
of in vivo and in vitro growing plantlets, aseptic seedlings, microspore mother cells, young
microspores, pollen grain calli and embryogenic and non-embryogenic suspension cultures.
More recently, viable protoplasts have been obtained from male and female gametes.
Direct method - In one step method, the leaf segments are incubated overnight (15-18h) with
enzyme mixture at 25°C and teased gently to liberate the protoplasts. The mixture is filtered
through fine wire gauze to remove leaf debris, transferred to 13 × 1000 mm screw capped tubes
and centrifuged at 100g for 1 min.
The protoplasts form a pellet and supernatant removed. The process is repeated three times
and protoplasts washed with 13% Sorbitol solution, which is later replaced by 20% sucrose
solution. The protoplasts suspension is centrifuged at a speed of 200g for 1 min. The cleaned
protoplasts, that are floating (debris settles down), can be pipetted out and bulked.
Sequential method - In two step method, leaf segments with mixture A (0.5% macerozyme +
0.3% potassium dextran sulphate in 13% mannitol at pH 5.8) are vacuum infiltrated for 5 min;
transferred to a water bath at 25°C and subjected to slow shaking.
After 15 min. the enzyme mixture is replaced by fresh ‘enzyme mixture A’ and leaf segments
incubated for another hour. The mixture is filtered using nylon mesh, centrifuged (100g) for 1
min. and washed three times with 13% mannitol to get a pure sample of isolated cells.
These cells are then incubated with ‘enzyme mixture B’ (2% cellulase in a 13% solution of
mannitol at pH 5.4) for above 90 min at 30°C. After incubation, the mixture is centrifuged at
100g for 1 min, so that protoplasts form a pellet, which is cleaned three times as in ‘one step
method’ above.
Factors affecting yield and viability of protoplasts
Source of material
Leaf has been the most favorite source of plant protoplasts because it allows the isolation of a
large number of relatively uniform cells without the necessity of killing the plants. Since the
mesophyll cell is loosely arranged, the enzymes have an easy access to the cell wall. The
leaves from in vitro roots or shoots released twice as many viable protoplasts as the leaves from
field grown material. Owing to the difficulty in isolating culturable protoplasts from leaf cells of
cereals and other species, their cultured cells have been used as an alternative source material.
Pre enzyme treatment
The lower epidermis is peeled and floats the stripped pieces of leaf on the enzyme in manner
that the peeled surface is in contact with the solution. This will facilitate easy penetration of
enzyme in to intercellular spaces of leaf. Mesophyll protoplasts of cereals could be isolated
within 2 hr by infiltrating the leaf pieces with enzyme solution under a partial vacuum for 3-5 min.
The criterion used to check adequate infiltration is that leaf pieces will sink when the vacuum is
removed. Brushing the leaf with a soft brush or with the cutting edge of scalpel may also
improve enzymatic action.
Enzyme treatment
The two enzymes, essential to isolate protoplasts from plant cells are cellulase and pectinase.
Pectinase degrades mainly the middle lamella and the cellulase required to digest the cellulosic
cell wall. The crude commercial enzymes carry nucleases and proteases as impurities which
may be harmful to protoplasts viability. The activity of the enzymes is pH dependant and it is
also affected by the temperature. The optimal temperature for the activity of these enzymes is
40-50C which happens to be too high for the cells. Generally 25-30C is found adequate for
isolation of protoplasts.
Osmoticum
A fundamental property of isolated protoplasts is their osmotic fragility and hence, there is a
need for a suitable osmotic stabilizer in the enzyme solution, the protoplast medium and the
protoplast culture medium. Protoplasts are more stable in a slightly hypertonic rather than
isotonic solution. a higher level of the osmoticum may prevent bursting and budding but it may
inhibit the division of the protoplast. The most widely used osmotica are sorbitol and mannitol in
the range of 450-800 mmol.
Culture of protoplasts
The methods used for protoplasts culture are basically the same as those employed for other
tissue and cell culture. Protoplasts can be cultured on liquid or solid agar media to meet special
requirements. In the following section, a wide range of available culture methods are described.
Culturing in liquid media
Liquid cultures
The protoplasts are suspended in a small volume of liquid culture medium at an appropriate
density and placed in petri dishes, which are then sealed with parafilm to reduce the loss of
water from the culture medium. The advantage of liquid culture is that it allows gradual change
of the osmolarity of the culture medium and in this way promotes rapid cell regeneration. This
method requires relatively large volumes of protoplast suspension. If small volumes of
protoplasts have to be cultured, one of the following methods can be adopted.
Drop cultures
Small droplets (40 to 100 ul) of protoplasts suspension are placed on the inner side of the lid of
a petri dish. When the lid is covered on the bottom, the culture drops are changed towards the
bottom dish. To the dish, fresh medium can be added in small drops when required.
Microchamber cultures
Microchamber cultures are similar to hanging drop cultures and are adopted for individual
protoplast culture. A drop of protoplast suspension is placed on sterile cover glass and inverted
on the slide with microchamber. Microchamber culture offers an optically better view since the
depth of the chamber is kept at minimum.
Multiple drop array technique
In drop culture technique, five to ten relatively large drops are placed on the petri dish. In the
multiple drop array technique, the drop is reduced to 40 ml so that 50 drops can placed in a
single petri dish. This method is used to screen a wide range of nutritional and hormonal factors.
Microdroplet cultures
Microdroplet culture is used to culture individual protoplasts. For this technique, the size of
drops is reduced to 0.25 to 0.50 ml so that each droplet contains only one protoplast and special
cups or petri dishes are used.
Culturing on semi-solid media
Agar as gelling agent
The protoplast suspension is mixed with equal volume of melted agar medium kept at about 43
to 45 [0] C. Actually the agar plating technique was originally used for the plating of cell suspension
cultures and the method was later modified and applied to protoplast culture.
Agarose or Alginate as gelling agent
Solidification of media with agarose instead of agar improves the protoplast culture efficiency.
The improved efficiency of the agarose may be due to the absence of contaminating substances
and neutrality. Protoplasts are plated in thin layers of agarose on top of already poured and
solidified media in petri dishes.
Combination of liquid and solid media
Gel embedded protoplast cultures
The protoplasts are incorporated into the whole medium before plating. The gelled agar or
agarose with protoplasts is then cut into several blocks, which are transferred to large volumes
of liquid culture medium and placed on shaker.
Semi-solid media for liquification
Semi-solid media prepared of agar or agarose are generally used. In this type of culture,
protoplasts are plated on semi-solid media. The semi-solid media with protoplasts are remelted
at 40oC for 1 or 2 hours to recover protoplasts for further multiplication.
Different feeder techniques
In protoplast culture, minimum-plating density (mpd) is an important factor. The mpd can be
maintained at low level by using the following techniques.
Feeder layers
A layer mixture of protoplasts of different species is plated on an agar-solidified medium. The
protoplasts are subject to irradiation to inactivate but not kill the layer of protoplasts. This layer is
called feeder layer. Then the protoplasts to be cultured can be plated at lower density of 5 - 50
protoplasts/cm [3] .
Nurse cultures
In nurse culture technique, protoplasts of one or more species grown on a medium support the
growth of other species i.e., protoplasts can be cultured on an established protoplast culture.
Generally this is followed to culture the fusion products of two different protoplasts.
Reservoir media
Protoplasts are cultured in quadrate plates. The liquid medium is placed in two quadrates and
protoplast suspension in other two quadrates of the plate. The continuous leakage of medium
keeps up the viability of protoplasts.
Use of filter paper discs
A filter paper disc is placed on an agar medium over which protoplast suspension is poured.
The filter paper provides a physical support to the protoplasts and absorbs unwanted toxic
substances.
Purification of protoplasts
After the material has been incubated in enzymes solution for an adequate period the
incubation vessel is gently swirled or the leaf pieces are gently squeezed to release the
protoplasts held in the original tissue. The digestion mixture consists of sub cellular debris,
especially chloroplasts, vascular elements, undigested cells and broken protoplasts besides
intact and healthy protoplasts. The protoplasts isolated as above are present in the media
together with a range of cell debris and broken cell organelles. A number of methods are
available for purification of protoplasts from this mixture. Only two commonly used methods are
briefly described.
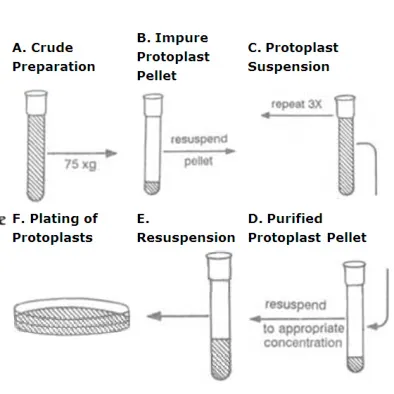
Sedimentation and washing
In this method, the crude protoplasts suspension is centrifuged at low speed (50-100g for 5
min). The intact protoplasts form a pellet and supernatant containing cell debris can be pipetted
off. The pellet is gently resuspended in fresh culture media plus mannitol and rewashed. This
process is repeated two or three times to get relatively clean protoplast preparation.
Protoplasts can be purified by repeated gentle pelleting and resuspension
Flotation
Protoplasts being lighter (low density) then other cell debris, gradients may be used, which will
allow the protoplasts to float and the cell debris to sediment.
A concentrated solution of mannitol, Sorbitol and sucrose (0.3-0.6M) can be used as a gradient
and crude protoplasts suspension may be centrifuged in this gradient at an appropriate speed.
Protoplasts can be pipetted off from the top of the tube after centrifugation.
This method causes little loss or damage relative to that in the ‘sedimentation and washing’
method. Babcock bottle is also used for flotation, since it facilitates removal of protoplasts.
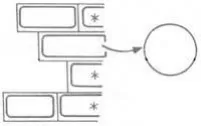
 Flotation of the protoplasts in a Babcock bottle greatly eases the removal of purified
Flotation of the protoplasts in a Babcock bottle greatly eases the removal of purified
protoplasts from the sucrose cushion
- Protoplasts, 2. Babcock bottle, 3. Flotation cushion
Viability of the protoplasts
Viability of the freshly isolated protoplasts can be checked by a number of methods:
Observation of cyclosis or cytoplasmic streaming as an indication of active metabolism.
Oxygen uptake measured by an oxygen electrode which indicated respiratory
metabolism.
Photosynthetic activity
Exclusion of Evan’s blue dye by intact membranes
Staining with fluroscein diacetate- which is most commonly used.
Protoplast culture
Protoplasts may be cultured in agar plates. An advantage in using semi-solid medium is that the
protoplasts remain stationary which makes it convenient to follow the development of specific
individuals. However, liquid medium has been generally preferred for the following reasons:
- The osmotic pressure of the medium can be effectively reduced after a few days of
culture
- Protoplasts of some species would not divide if plated in agarified medium.
 3. If the degenerating component of the protoplast population produces some toxic
3. If the degenerating component of the protoplast population produces some toxic
substances which could kill the healthy cells it is possible to change the medium.
- The density of cells can be reduced or cells of special interest may be isolated after
culturing them for a few days at a high density.
The protoplasts suspension is plated as a thin layer in petriplates, or incubated as static cultures
in flasks or distributed in 50-100 l drops in petri plates and stored in a humidified chamber.
Embedding protoplasts in agarose beads or discs is reported to improve plating and
regeneration efficiency in many species. Alginate is another gelling agent used for culture of
protoplasts, particularly of the species, which are heat sensitive such as Arabidopsis thaliana.
After 2-4 days in culture, protoplasts lose their characteristic spherical shape and this has been
taken as an indication of new wall regeneration. While the presence of a proper wall is essential
for regular division, not all such cells regenerated from protoplasts embark upon division. In
protoplast cultures, the cell divisions are asynchronous. The first division may be equal or
unequal. Mitosis is normal. Continuous cell division leads to callus formation and the plant will
be regenerated through normal developmental process.
Plant regeneration
Protoplasts, thus cultured undergo the following processes to produce plantlets. They are:
- Cell wall formation
- Cell division and callus formation
- Plant regeneration
Cell wall formation
Protoplasts in culture start to regenerate cell wall within a few hours, and may take two to
several days to complete it. Within 2-4 days in culture, protoplasts lose their characteristic
spherical shape and this has been taken as an indication of new wall regeneration. The wall
synthesis by protoplasts starts immediately after the enzyme is washed off. During cell wall
formation, the cellulose is deposited either between plasmalemma and multilamellar wall
material or directly on the plasmalemma. A freshly formed cell wall is composed of loosely
arranged microfibrils, which subsequently become organised to form a typical cell wall. The
protoplasts may start cell wall synthesis 10-20 minutes after culture or go without cell wall for
over a period of seven days. The protoplasts with normal cell wall undergo mitosis and produce
daughter cells. The protoplasts with poorly formed cell walls do not undergo normal mitosis, but
fuse with each other to produce multinucleate cells or enlarge in size to undergo budding.
Cell division and callus formation
Normally after cell wall regeneration, the cell undergoes a significant increase in size. This is
followed by first mitotic division. Immediately after first division, the protoplasts may undergo a
lag phase, which lasts for 7-25 days. Generally, protoplasts of actively growing cell suspensions
undergo first division faster than those from mesophyll protoplasts. The second round of
divisions is often observed within a week of the first division. Small cell clumps form within two
weeks of second division producing small pieces of callus.
Plant regeneration
The general techniques applicable to plant regeneration from tissue cultures hold good for the
callus obtained from protoplasts also. The first step for the regeneration of plants involved the
transference of callus to regeneration medium containing balanced phytohormones either to
induce organogenesis or somatic embryogenesis. The first report of plant regeneration from
isolated protoplasts was from Nicotiana tabacum by Takebe et al ., in 1971. Since then the list of
species exhibiting this potentiality had steadily increased.
Applications
Virus uptake: Studies on the mechanism of infection and host parasite relationships
Bacterial uptake: Symbiotic nitrogen fixing bacterium ( Rhizobium, Azotobacter ) can be
introduced into legume. Direct DNA transfer and expression of a bacterial gene in protoplasts of
exogenous DNA by cells or protoplasts of T. Monococcum and N. tabucum are reported.
Incorporation of Cyanobacterial cells (e.g.): Cyanobacteria or BGA. Co-incubate algal
preparation with isolated protoplasts with 25% PEG and high planting density. Protoplasts begin
engulf algal cells.
Incorporation of exogenous DNA: Exogenous DNA can be taken up by higher plant
cells/protoplasts and this is known as Trasngenosis.
Transplantation of nuclei: Organelles such as large nuclei can be introduced through plasma
lemma into protoplasts. Both intra and inter specific nuclear transplantations have been
observed in Petunia hybrida, Nicotiana tabacum and Zea mays .
Protoplast fusion
The feature of isolated protoplasts that has brought them into the limelight is the ability of these
naked cells to fuse with each other irrespective of their origin. The technique of hybrid
production through the fusion of body cells, bypassing sex altogether, is called somatic
hybridization. Unlike sexual reproduction in which organelle genomes are generally contributed
by the maternal parent, somatic hybridization also combines cytoplasmic organelles from both
the parents. In somatic hybrids, recombination of mitochondrial genome occurs frequently.
Chloroplast genome recombination is rare but segregation of chloroplasts of one or the other
parent, forming novel nuclear-cytoplasmic combinations. Fusion products with the nucleus of
one parent and extra-nuclear genome/s of the other parent are referred to as cybrid and the
process to obtain cells or plants with such genetic combination/s are called cybridization.
During enzymatic degradation of cell walls some of the adjacent protoplasts fuse together
forming homokaryones (also referred as homokaryotes each with two to several nuclei). This
type of protoplast fusion is called as spontaneous fusion. A sequential method of protoplast
isolation or exposing the cells to strong plasmolyticum solution before treating them with mixed
enzyme solution would affect the plasmodesmatal connection and consequently reduce the
frequency of spontaneous fusion. So far as somatic hybridization concerned spontaneous fusion
is of no value; these require the fusion of protoplasts of different origin. To achieve induced
fusion, a suitable chemical agent (fusogen) or electric stimulus is generally necessary.
Chemical fusion
NaNO3 treatment
Hypotonic solution of NaNo3 induces the fusion of sub-protoplasts within plasmolysed
epidermal cells. However, this technique suffers from a low frequency of heterokaryon
formation.
High pH and high Ca [2+] treatment
Mesophyll protoplasts of two different lines of the species could be fused by treating them in a
highly alkaline solution (pH 10.5) of high Ca [2+] ions at 37C for about 30 min. intra and inter
specific Nicotiana hybrids were developed using this technique. However, for some species this
high pH may be toxic.
Polyethylene glycol (PEG) treatment
PEG has accepted as fusogen because of the reproducible high frequency heterokaryon
formation, especially binucleate formation, and its comparatively low cytotoxicity to most cell
types. PEG induced fusion is non-specific. In addition to fusing soybean-maize and soybean
barley, PEG brings about effective fusion between animal cells, animal cells with yeast
protoplast and animal cells with higher plant protoplasts. The isolated protoplasts of the two
selected parents are mixed in appropriate proportions and treated with 15-45% PEG (1500
6000 MW) solution for 15-30 min followed by gradual washing of the protoplasts with the culture
medium.
Mechanism of protoplast fusion
Protoplast fusion consists of three main phases:
- Agglutination, during which the plasma membrane of two or more protoplasts are brought into
close proximity.
- Membrane fusion at small localized regions of close adhesion resulting in the formation of
cytoplasmic continuities or bridges between protoplasts.
- Rounding off of the fused protoplast due to the expansion of the cytoplasmic bridges forming
spherical hetero- or homokaryons.
The actual mechanism of PEG-induced fusion is not clear. The accepted hypothesis is that the
PEG molecule which is slightly negative in polarity can form hydrogen bonds with water, protein,
carbohydrate etc., which possess positively polarized groups. When the PEG molecule chain is
large enough it acts as a molecular bridge between the surface of adjacent protoplasts and
adhesion occurs. PEG can bind Ca [2+] as well as other cations. These calcium ions may form a
bridge between the negatively polarized groups of protein (or phospholipids) and PEG, thus,
enhancing adhesion. During the washing process the PEG molecules bound to the membranes
either directly or through Ca [2+] is eluted, resulting in disturbance and redistribution of the electric
charge. Such a redistribution of charge in the regions of intimate contact of the membranes can
link some of the positively charged groups of one protoplast to the negatively charged groups of
other protoplast and vice versa, resulting in protoplast fusion.
Disadvantages of chemical fusion
-
The fusogens are toxic to some cell types.
-
It produces random, multiple cell aggregates.
Electro fusion of protoplasts
Electro fusion is rapid (usually completes within 15 min), simple, synchronous and more easily
controlled. It was shown that electrofusion is more effective than PEG mediated fusion in
somatic hybridization of Solanum tuberosum and S. brevidens. Zimmermann et al., (1982)
developed a method, Zimmermann Electrofusion System, which is claimed to be 10000 times
better than any other method for protoplast fusion.
This technique utilizes low voltage electric current pulses to align the protoplast in a single row
like a pearl-chain. The aligned protoplasts are pushed, with a micromanipulator, at a gentle
pace through the narrow gap between the two electrodes. When the two protoplasts that are to
be fused are appropriately oriented opposite the electrodes, a short pulse of high voltage is
released which induces the protoplasts to fuse. The high voltage creates transient disturbances
in the organization of plasmalemma, which leads to the fusion of neighbouring protoplasts. The
entire operator is carried out manually in specially devised equipment, called electroporator,
under a microscope.
Selection of hybrid cells
The protoplast suspension recovered after a treatment with a fusion-inducing agent consists of
the following cell types:
-
Unfused protoplasts of the two parents
-
Products of fusion between two or more protoplasts produced by fusion between the same
parents (homokaryon)
- Hybrid protoplasts produced by fusion between one or more protoplasts of each of the two
species (heterokaryon). Heterokaryon particularly those resulting from one protoplast of each of
the two parents are of interest; these forms usually in a very small proportion (0.5 –10 %).
These hybrid protoplasts are identified by the following strategies.
Visual markers
e. g. pigmentation of the parental protoplasts. The protoplast of one parent may be green and
vacuolated (mesophyll cells), while those of the other may be non-vacuolated and nongreen
(from cell cultures). If they are not having these features, then these protoplasts are differently
labeled with fluorescent dyes. However, this approach is time consuming and requires
considerable skill and effort.
Complementation
The property, which is not present in one parent, will be acquired when they are in hybrid state.
For example, protoplasts of Petunia hybrida form calli on the MS medium, while those of P.
parodii produce only small cell colonies. Further, actinomycin D inhibits cell division of P.
hybrida protoplasts, but it has no effect on those of P. parodii. Thus protoplasts of both the
species fail to produce macroscopic colonies on MS medium supplemented with actinomycin D.
However, their hybrid cells ( Petunia hybrida + P. parodii) (note this symbol to denote the
somatic hybrids) divide normally on this medium to produce macroscopic colonies. These
strategies are simple, highly effective and least demanding. But their applicability is drastically
affected by the non availability of suitable properties in most of parental species.
Culture the entire protoplast population
The entire protoplast population is cultured without applying any selection for hybrid cells. All the
types of protoplasts for calli; the hybrid calli are later identified on the basis of callus
morphology, chromosome constitution, protein and enzyme banding pattern etc.
Cybridization
In sexual hybridization the plastid and mitochondrial genomes are generally contributed by only
the female parent whereas in somatic hybridization the extra nuclear genomes from both the
parents are combined. Cybrids or cytoplasmic hybrids are cells or plants containing nucleus of
one species but cytoplasm from both the parental species. They are produced in variable
frequencies in normal protoplast fusion experiments due to one of the following:
- Fusion of a normal protoplast of one species with an enucleate protoplast or a protoplast
having an inactivated nucleus of the other species
-
Elimination of the nucleus of one species from a normal heterokaryon
-
Gradual elimination of the chromosomes of one species from a hybrid cell during the
subsequent mitotic divisions.
Cybrids may be produced in relatively high frequency by
- Irradiating (with X or Gamma rays) the protoplasts of one species prior to fusion in order
to inactivate their nuclei
- By preparing enucleate protoplasts (cytoplasts) of one species and fusing them with
normal protoplasts of the other species.
The objective of the cybrid production is to combine the cytoplasmic genes of one species with
the nuclear and cytoplasmic genes of another species. But the mitotic segregation of plasma
genes, as evidenced by the distribution of chloroplasts, leads to the recovery of plants having
plasma genes of one or the other species only. Only a small proportion of the plants remain
cybrid.
This provides the following applications:
- Transfer of plasma genes of one species into the nuclear background of another species
in a single generation and even in sexually incompatible species.
-
Recovery of recombinants between the parental mitochondria or chloroplast DNAs.
-
Production of wide variety of combinations of the parental and recombinant chloroplasts
with the parental or recombinant mitochondria.
- Mitochondria from one parental species and chloroplast from another parental species
may be combined.
The cybrid approach has been used for the transfer of cytoplasmic male sterility from
Nicotiana tabacum to N. sylvestris and P. hybrida to P. axillaries.
Advantages of production of CMS lines through cybridization
Only one step is required.
The nuclear genotype of cultivar remains unaffected.
100% of the progenies of somatic hybrids will be CMS.
Questions
- The feasibility of enzymatic degradation of plant cell walls to obtain large quantities of
protoplasts was demonstrated by …………
a). E. C. Cocking b). Klecker c). Takebe d). None of the above
- The protoplasts were isolated by …………
a). Mechanical method b). Enzymatic method c). Both a and b d). None of the above
- The protoplast isolation by enzymatic method was demonstrated by …………
a). E. C. Cocking b). Klecker c). Takebe d). None of the above
- The enzyme used in enzymatic method for protoplast isolation is …………
a). Cellulase b). Pectinase c). Both a and b d). None of the above
- The commercial preparations of the enzymes for protoplast isolation were first employed
by …………
a). E. C. Cocking b). Klecker c). Takebe et al d). None of the above
- The commercial preparation of the enzymes for protoplast isolation was first employed by
…………
a). E. C. Cocking b). Klecker c). Takebe et al d). None of the above
- The leaf has been the most favorite source of plant protoplasts isolation because
…………
a). isolation of a large number of relatively uniform cells without the necessity of killing the plants b). mesophyll cell is loosely arranged and the enzymes have an easy access to the cell wall c). both a and b d). None of the above
- The most widely used osmotica for protoplast isolation was/were…………
a). sorbitol b). mannitol c). both a and b d). None of the above
- The culturing of protoplast in liquid media includes …………
a). Drop cultures b). Microchamber cultures c). Multiple drop array technique and d). All the above microdoplet cultures
- The culturing of single protoplast in liquid media is done by …………
a). Drop culture method b). Microchamber culture c). Multiple drop array technique d). Microdoplet cultures
- The minimum-plating density (mpd) in protoplast culture is maintained by …………
a). Feeder layers b). Nurse cultures c). Reservoir media d). All the above
- The first report of plant regeneration from isolated protoplasts was from …………
a). Nicotiana tabacum b). Petunia hybrida c). Z ea mays d). None of the above
- The first report of plant regeneration from isolated protoplasts was by …………
a). Takebe b). Klecker c). E. C. Cocking d). None of the above
- For protoplast fusion ………… is/are used
a). NaNO3 b). High pH and high Ca [2+] c). PEG d). All the above
- The mechanism of chemical fusion of protoplast consists of …………
a). Agglutination b). Membrane fusion c). Rounding off of the fused protoplast d). All the above
- The electro fusion of protoplast is …………
a). Rapid and simple b). Synchronous c). More easily controlled d). All the above
Summary Cheat Sheet
Quick Recall Points
- A protoplast is a plant cell without its cell wall.
- E. C. Cocking demonstrated enzymatic protoplast isolation in 1960.
- Mechanical isolation is historically important, but enzymatic isolation is the more practical method.
- Protoplast fusion is used in somatic hybridization to overcome sexual incompatibility barriers.
Exam Traps
- Do not confuse protoplast isolation with protoplast fusion; they are related but different steps.
- Naked cells are obtained only after proper wall removal and osmotic protection.
- Fusion products may show chromosome imbalance, so not every hybrid is a stable amphiploid.
References
1 source • [1]
References
Standard BSc Agriculture Plant Biotechnology notes
BookLesson Doubts
Ask questions, get expert answers